Research Article - Annals of Biological Research ( 2018) Volume 9, Issue 3
Yield is an economically important trait considered by farmers engaged in taro production. The objective of the present study was to increase genetic diversity and assess the yield stability performance of the new taro cultivars evaluated under different environments. Fourteen cultivars were evaluated for two years 2013-2014 and 2015-2016 including other cultivars. The trial was planted in an augmented design and data was pooled and analyse as a randomised complete block design. The corm yields analysed using the additive main effect and multiplicative interaction (AMMI) and genotype and genotype by environment interaction (GGE) biplot. The additive main effects and multiplicative interaction model (AMMI) analysis of variance showed significant differences for genotype, environment and genotype by environment interaction. The result showed 31% of the total variation was accounted by environment while genotype and interaction accounted for 16.6 and 26.4%, showing significant differences. The highest variation by environment, (GXE) indicates that the genotypes performed differently across environments. The C5-353 was the ideal genotype with both high yield and high stability as indicated by the AMMI stability value (ASV) analysis and yield stability index (YSI) showing wider adaptability. The other most stable genotypes were BL/SM/43 and BL/SM/148. The GGE biplot indicated that M02 was the most stable environment and C5-353 was the most stable cultivar.
Genotype by environment, Main effect and multiplicative interaction (AMMI), Biplot, Genotype, Stability, Genetic diversity
Taro (Colocasia esculenta) is an essential horticultural crop in the tropics and is deep rooted in the culture of Pacific people in many traditional societies [1,2]. The breeding efforts to improve taro production in Papua New Guinea has been rather slow since the start of the breeding program in 1993. The current breeding program is based on recurrent selection method aim at improving resistant to taro leaf blight (TLB) [3]. To continue breeding, continued success can only be achieved through availability of diverse new genetic material to respond to narrow genetic diversity experienced by farmers and better statistical analysis to select potential cultivars or genotypes [4]. Though, it is rich in carbohydrate, vitamin nutrients (10) the total production remains low globally and in Papua New Guinea it is estimated at 274,620 t/ha [5].
Genotype by environment (GxE) yield trials are important to identify and select adaptable high yielding cultivars and discover sites that best represent the target environment [6]. The adaptability of a cultivar is a combination of genotype, environment and genotype by environment interaction. In this aspect, the cultivars can be identified to be widely adaptable or ability to perform best only in desirable environment, known as specific adaptability [7]. To be able to identify most adaptable varieties, this requires better statistical analysis that can captures both genotypic and environmental effects.
In this study, we employ the use of main effect and multiplicative interaction (AMMI) and genotype and genotype by environment interaction (GGE) biplot model. The two statistical models were used determined the pattern of genotypic responses across environments (GxE) [6,8]. The most commonly used statistical technique was the twoway analysis of variance (ANOVA). However, some breeders reported that anova lacks clear explanation on the interaction effect (GEI) [9]. Both AMMI and GGE biplot are useful as they uses a principal component to interpret genotype performance by integrating the use of ANOVA and principal component analysis (PCA). However, (AMMI) also incorporates additive components in a single model for the main effects of genotype and environment as well as multiplicative components for the interaction effect.
The GGEbiplot model on the other hand, uses graphic axes by identifying superior genotypes in the mega environments and integrating the ANOVA and PCA sum of squares of genotypes and genotype by environment using principle component (PCA) method. One of the advantages that GGE biplot has over the AMMI model is that which won where pattern is easily visualised in the GGE biplot.
Therefore, the objective of the current study was to identify high yielding and stable performing genotypes under different agro-ecologies using AMMI and GGE biplot analysis models to recommend for release in the test environments and other similar agro-ecologies in the country.
Planting materials and study site
Thirteen (11 cultivars & 2 advance breeding lines) sourced under the International Edible Aroids Network (INEA) and National Agricultural Research Institute (NARI). The cultivars and the advance breeding line were kept at the institutes germplasm collection in Lae, Papua New Guinea and one local commercial variety Numokowe (used as check) were evaluated across five different environments namely; Murukanam (M02) Murukanam (M02), Kerevat (K03), Yonki (Y04) and Salamaua (S05) (Table 1) and the morphological features are presented in Table 2. These trials were evaluated across four different provinces between 2014-2015. The trials were planted in an augmented design, the number of farmers per sites was used as replicates, and data rearranged as a randomized complete block design (RCBD) for analysis. Important agronomic traits, plant height, sucker numbers, stolon, flowering and taro leaf blight recorded and eating quality taste test conducted. However, only taro corm yield trait was used for stability data analysis.
| Environments | Province | Altitude (masl) | Rainfall (mm) | Soil texture | Soil type |
|---|---|---|---|---|---|
| Derin (D01) | Madang | <600 | 900-2800 | Black loam | Tropudalfs |
| Murukanam (M02) | Madang | <600 | 900-2800 | Sandy loam | Tropofluvents |
| Salamaua (S05) | Morobe | <600 | 2000-2500 | Sandy loam | Fluvaquents |
| Kerevat(K03) | East New Britain | <600 | 2500-3000 | Sandy-black loam | Tropofluvents |
| Yonki (Y04) | Eastern Highlands | <1500 | Black-clay loam | Aquolls |
Table 1: The description of the environments used in the study.
| Genotype | Origin | Type | Corm Flesh Colour | Corm Shape | Corm Branch | Corm Skin Surface |
|---|---|---|---|---|---|---|
| BL/SM/143 | Samoa | Breeders line | white | elliptical conical | unbranched | smooth |
| BL/SM/148 | Samoa | Breeders line | light yellow | conical | unbranched | smooth |
| BL/SM/43 | Samoa | Breeders line | pink | conical | unbranched | smooth |
| BL/SM/80 | Samoa | Breeders line | white | cylindrical | unbranched | smooth |
| C5-245 | PNG | Breeders line | purple | ellipticall | unbranched | smooth |
| C5-353 | PNG | Breeders line | white | ellipticall | unbranched | smooth with thick covering |
| BL/SM/104 | Samoa | Breeders line | light yellow | conical | unbranched | smooth |
| BL/SM/115 | Samoa | Breeders line | creamy white | conical | unbranched | smooth |
| BL/SM/128 | Samoa | Breeders line | pale yellow | elliptical | unbranched | smooth |
| BL/SM/136 | Samoa | Breeders line | purple | elliptical, conical | branched/unbranched | smooth |
| BL/SM/149 | Samoa | Breeders line | light yellow | elliptical, conical | unbranched | smooth |
| CE/IND/16 | Indonesia | Breeders line | yellow | cylindrical | branched | fibrous |
| CE/MAL/07 | Malaysia | Breeders line | light yellow | conical | branched | smooth |
| Numkowe | PNG | landrace | pink | ellipticall | unbranched | smooth |
Table 2: Genotype , origin and characteristics used in the study.
AMMI analysis
AMMI analysis was carried out for corm yield 11 taro cultivars and two advance breeding line including the check variety obtained per plot across environments using the Genstat software Release 14 edition. The AMMI model combines the analysis of variance for main effects of G and E with principal components analysis of GEI. It has proven useful in understanding complex genotype by environment interaction (GEI). Stable genotypes for each environment were selected by AMMI and the AMMI anova table and AMMI first four selection and principal component axes (PCAs) were extracted and statistically tested [10]. The AMMI statistical model used to combine the analysis of variance over environments was:
Where yger was the observed yield of genotype (g) in environment e for replication(r). The additive parameters (μ) was the grand mean, αg is the deviation of genotype g from the grand mean. βe is the deviation of the environment e and the multiplicative parameters: was the singular value for interaction principal components axis (IPCA) , Ygn was the genotype eigenvector for axis , and e is the environment eigenvector, ge are PCA residuals and ger is the error term . Accordingly, genotypes with low (regardless of the sign) IPCA scores showed general or wider adaptability, while those with large PCA scores shows specific adaptability [8,11].
AMMI stability value analysis (ASV)
The AMMI stability value (ASV) is defined as the distance from the co-ordinate point to the origin in a two dimensional plot of IPCA1 scores versus IPCA2 scores in the AMMI model [12]. Since the IPCA1 scores contributes more to the genotype by environment interactions sum of squares, a weighted value is needed. This was calculated for individual genotypes and environment according to the relative contribution of IPCA1 to IPCA2 as follows:

The larger the ASV value, either negative or positive the more specifically adapted a genotype was to certain environments [11]. A small ASV value indicated a more stable genotype across environments [12].
Yield Stability Index (YSI)
The yield stability index (YSI) was calculated as follows:YSI= RASV+RMG where RASV is the ranking of the AMMI stability value and RMG is the rank mean of the genotypes across all environments. YSI of low value shows high yield stability.
GGE biplot analysis
The genotype and genotype by environment analysis was carried using GGE biplot method outlined [6] in genstat 14 edition. The which won where pattern was visually presented using the GGE biplot inner product property of the biplot. Additionally, the GGE biplot stability analysis was performed to identify high yielding and adapted genotypes across test environments for possible release. The best taro genotypes were represented by large principal component scores (PCA 1, high corm yield) and small principal component scores (PCA 2, high stability) [13]. Genotypes that had PCA 1 scores >0 were identified as higher yielding and those that had PCA 1 scores <0 were identified as lower yielding [14]. The model for the GGE biplot based on singular value decomposition (SVD) of first two principal components based on [15,16].
where Yij is the mean of genotype I in environment j, μ is the grand mean. βj is the main effect of environment j, μ +βj is the mean yield across all genotypes in environment j. 1and 2 are the singular values (SV) for the first and second principle components (PCA1 and PCA2), i1 and i2 are the eigenvectors of genotype I for PCA and PCA2 respectively, 1j and 2j are eigenvectors of environment j.
The combined analysis of variance (ANOVA) for the yield across the five different locations are presented in Table 3. There was only highly significant difference in yield (P<0.05) in yield at different sites (Table 2). The overall yield means ranged from 7.1 t/ha to 14.9 t/ha, for C5-353, being the best while BL/SM/149, CE/MAL/07 and CE/IND/16 were the least.
| Genotype | D01 | M02 | K03 | Y04 | S05 | Cultivar mean |
|---|---|---|---|---|---|---|
| BL/SM/104 | 12.9 | 10.9 | 4.6 | 2.8 | 8.9 | 8 |
| BL/SM/115 | 10.9 | 16.1 | 6.1 | 9.2 | 13.5 | 11.1 |
| BL/SM/128 | 11.6 | 8.7 | 6.3 | 6 | 5.5 | 7.6 |
| BL/SM/136 | 16 | 10.6 | 7.9 | 1.8 | 7.8 | 8.8 |
| BL/SM/143 | 10.1 | 18.6 | 11.5 | 9.8 | 13.1 | 12.6 |
| BL/SM/148 | 21 | 14.3 | 7.1 | 6.4 | 7.3 | 11.2 |
| BL/SM/149 | 12.3 | 11.2 | 2.6 | 4.7 | 4.8 | 7.1 |
| BL/SM/43 | 15.6 | 18.2 | 7.4 | 4.7 | 8.7 | 10.9 |
| BL/SM/80 | 16.8 | 9.9 | 7.2 | 3.2 | 7.7 | 8.9 |
| C5-245 | 14.3 | 12.6 | 4.8 | 14.2 | 9.2 | 11 |
| C5-353 | 21.1 | 24.1 | 9.4 | 12.3 | 7.7 | 14.9 |
| CE/IND/16 | 7.9 | 10.1 | 4.3 | 11.9 | 4.4 | 7.7 |
| CE/M AL/07 | 7.3 | 9.7 | 6.8 | 8.17 | 5.6 | 7.5 |
| NUMKOWE | 7.6 | 14.3 | 7.6 | 8.7 | 9.1 | 9.5 |
| Environment mean | 13.2 | 13.5 | 8.9 | 7.4 | 6.6 | 9.7 |
| CV% | 20.7 | 37.6 | 37.3 | 30.7 | 34.3 | - |
| P<(0.05) | 0.002 | 0.24 | 0.2 | 0.06 | 0.13 | - |
Table 3: Mean corn yield (t/ha) of the cultivars across five different environments.
Genotype by environment interactions were significant (P<0.05) for the corm yield trait (Table 4). In terms of the environments there was highly significant differences observed only in environment (D01) with exception to other four environments (Table 3) which had low mean average yields. The highest yields were recorded in D01 and M02 but were similar (13.2 to 13.5 t/ha) respectively (Table 3).
Specific selection for individual environments show that C5-353 and C5-245 are high potential genotypes for higher altitude growing regions of the country (Y04) (Table 4). The following cultivars are potential genotypes for lowland growing environments (D01, M02, K03 and S05) are C5-353, BL/SM/143,148,115 and BL/SM/80 (Table 4). The C5-353 is considered a genotype that can be widely adapted to both lowland and highlands conditions.
AMMI gxe and stability analysis
AMMI analysis of variance showed highly significant (p<0.05) effects of genotype, environment and GxE interaction (Table 4). The AMMI analysis of the taro corm yield for the cultivars evaluated across five different environments showed that the largest variation (31%) was accounted by environment, GxE interaction effect accounted for (26.4%). The genotypes contributed only 16.6% of the total variation. The large sum of squares by environment shows that the environment in which the trials were conducted were diverse with differences among environmental means contributing to the variation in corm yield (t/ha). Total treatment variation accounted for 74% and 6.7% is residuals. This implies that different varieties could be selected for the different environments (Tables 4 and 5). The significant GxE interaction of corm yields implies that different genotypes perform differently across the five environments and the large sums of squares shows large variation amongst the environments over cultivars (Table 5). From this study, the use of the AMMI model was justified and it effectively partitioned the sum of squares (Table 5). Similar results were reported by Farshadfar et al. [17] in their studies on 20 wheat genotypes evaluated under two different environments for years showing significant variations among genotypes environment and environment GE interactions which were also reported in bread wheat, rice, finger millet, common bean 12). IPCA 1 followed by IPCA 2 explained a substantial amount of percentage of GxE interaction. Several authors took the first two IPCA for AMMI analysis because the greater percentage of genotype by environment interaction (GxE) in most cases was explain by first IPCA scores such as for bread wheat [17,18] and finger millet [11].
| Environment | NE | IPCA 1 | IPCA2 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|---|---|
| D01 | 1 | -3.21634 | -1.0542 | C5-353 | BL/SM/148 | BL/SM/43 | BL/SM/80 |
| M02 | 2 | 0.0105 | -0.14358 | C5-353 | BL/SM/143 | BL/SM/148 | C5-245 |
| K03 | 3 | 0.1419 | 1.20975 | BL/SM/143 | C5-353 | BL/SM/115 | BL/SM/43 |
| Y04 | 4 | 2.39938 | -2.04799 | C5-353 | C5-245 | CE/IND/16 | BL/SM/143 |
| S05 | 5 | 0.66457 | 2.03603 | BL/SM/143 | BL/SM/115 | NUMKOWE | BL/SM/43 |
Table 4: The first four selections by AMMI analysis.
AMMI stability value (ASV) and yield stability index (YSI)
With the AMMI model, one disadvantage is that it lacks the capability of making provision for a quantitative stability measure, such a measure is important in order to rank cultivars according to their yield stability. Therefore, we adopted the use of AMMI stability value (ASV) to cope with this problem. The ASV is in fact the distance from zero in a two-dimensional scatter graph of IPCA1 (interaction principal component analysis axis 1) scores against IPCA2 scores. The distance from zero is then determined and in an ASV method, a genotype with least ASV score is the most stable. An ideal genotype should have high corm yield and small ASV value. Accordingly, genotype BL/SM/ 128, and 43 from Samoa showed the lowest ASV score (0.00, 1.18) and moderate corm yield (7.6, 10.9t/ha). Furthermore, Samoan line BL/SM/115 showed high yield with a relatively low ASV followed by BL/SM/104. The result shows that these genotypes are showing relatively better stability than other genotypes. However, stability needs to be considered in combination with yield [17]. Thus, PNG advance breeding line C5-353, C5-245 and Samoan lines 143 and 148 were the highest yielding genotypes (14.9, 12.6,11.0, 11.2, t/ha respectively with high ASV 3.45, 2.82, 3.54,3.72 respectively. These genotypes were identified as good genotypes to validate for yield performance and specific adaptability with similar studies reported on finger millet [11]. The results of the ASV confirms that CE/IND/16 was unstable and CE/MAL/07 was consistently producing low yield across environments. Furthermore, [17] reported on 20 bread wheat genotypes evaluated gave small ASV and higher grain yield than the grand mean and stability.
Additionally, ASV is not the only parameter for selection of a new genotype. Considering this, the most stable genotypes would not necessarily give the best yield performance or may give higher ASV value. Hence, there is a need for alternative approaches that incorporate both mean yield and stability in a single index. Therefore, we applied the yield stability index (YSI) based on the rank of ASV and yield mean. This analysis was used in such a way that the lowest ASV takes the rank one, while the highest yield mean takes the rank one and then the ranks are summed in a single simultaneous selection index of yield to produce yield that is stable (Table 4). The least YSI is considered as the most stable with high corm yield. Based on YSI the most stable genotypes were identified and selected BL/SM/ 148, 143, 43, C5-353, and C5-245, these results are in accordance with the rank mean (Table 5).
| Source | df | SS | MS | Variation (%) |
|---|---|---|---|---|
| Total | 139 | 4005 | 28.8 | |
| Treatments | 69 | 2964 | 42.9*** | 74 |
| Genotypes | 13 | 665 | 51.2*** | 16.6 |
| Environments | 4 | 1241 | 310.3*** | 30.9 |
| Block | 5 | 265 | 53.1*** | 6.6 |
| GxE | 52 | 1057 | 20.34** | 26.4 |
| IPCA1 | 16 | 549 | 34.29*** | 13.7 |
| IPCA2 | 14 | 239 | 17.1ns | 5.9 |
| Residuals | 22 | 270 | 12.25 | 6.7 |
| Error | 65 | 776 | 11.93 |
Table 5: Combined analysis of variance in taro varieties across five different environments ns- not significant,** significant at 5% *** highly significant at 5%,
When running the AMMI analysis, we concentrated our analysis on the test genotypes and the commercial local cultivar Numkowe. From the AMMI analysis, the AMMI analysis selects genotypes PNG advance breeding line C5-353, BL/SM/143, BL/SM/43 and BL/SM/148 performed absolutely well in all five environments (Table 3). However, the differences in ranking of these genotypes in the different environments implied differential yield performance among the taro genotypes because of the significant genotype by environment interactions. According to Farshadfar et al and Nzuve et al. [19,20] this is also referred to as crossover effect of the GEI. The GGEbiplot and the AMMI Biplot ensure environment stratification while defining cultivars which optimise performance in the environment [21].
The corm yield adaptability and stability using GGEbiplot analysis
The corm yield adaptability and yield stability of the different taro genotypes were visually assess graphically using the GGE biplot analysis. In our study the two principle components explained a total of 78 % GGE variation in which PC1 accounted for 46.66% and PC2 31.17%, Figure 1). The PCA1 was used to estimate the yield in that genotypes, which had higher PC1 scores, considered more productive. The second principle component which was represent along the y axis explained the stability of the genotypes [20,22]. The average environment axis (AEA) or sometimes known as average tester coordinate (ATC) was presented as the line which pass through the origin of the biplot indicated by an arrow head. In our study (Figure 1) further shows the stability of the genotypes across the test environments. Cultivars C5-353 had shortest projection length and is highly stable followed by cultivars BL/SM/43, C5-245. The rank of the cultivars from high to low in terms of their yield according to the stability analysis is C5-353, BL/SM/148, BL/SM/143, BL/SM/43, BL/SM/115 and C5-245. The PNG advance breeding line C5-353 genotype could be used for broad selection because it was found to be the most ideal genotype with both high mean yield and high stability across environments. We confirm that GGE biplot can be used to select highly stable cultivars. Our findings are similar to a study conducted by Sharma et al. [23] on identifying high yielding stable winter genotypes under irrigated environments.

Figure 1: Mean performance and stability of the taro 13 genotypes using GGE biplot analysis.
GGE-biplot analysis of yield using PC1 and PC2 (Figures 2 and 3). This Figure 4 shows the grouping of the genotypes according to high yielding or low yielding by average ordinate environment (AOE). The dotted line shows the performance line, which passes through the origin of the biplot that helps to determine the mean performance of a genotype. This figure shows which genotypes performs best where and which is the best for an environment. Therefore, genotypes C5-353, BL/SM/148, BL/SM/143, BL/SM/43, and BL/SM/115 were identified as the best performing genotypes. Besides this, other genotypes that had large negative PC1 scores and low yield (Figure 1) were genotypes CE/MAL/07, CE/IND/16, BL/SM/128 and BL/SM/149, BL/SM/136. Genotypes C5-353, and BL/SM/43 are more stable followed by BL/SM/148 and BL/SM/143. Among these genotypes, C5-353 had the highest yield (t/ha). The genotypes BL/SM/143, C5-353 and BL/SM/148 were identified as the best performing genotypes. From this study, the genotypes C5-353, BL/SM/148, C5-245, BL/SM/143, 43 and 115 showed the best performance in M02 and D01 and Y04 environments respectively. They were also the most responsive genotypes amongst the check which showed specific adaptations and wider adaptations [22].
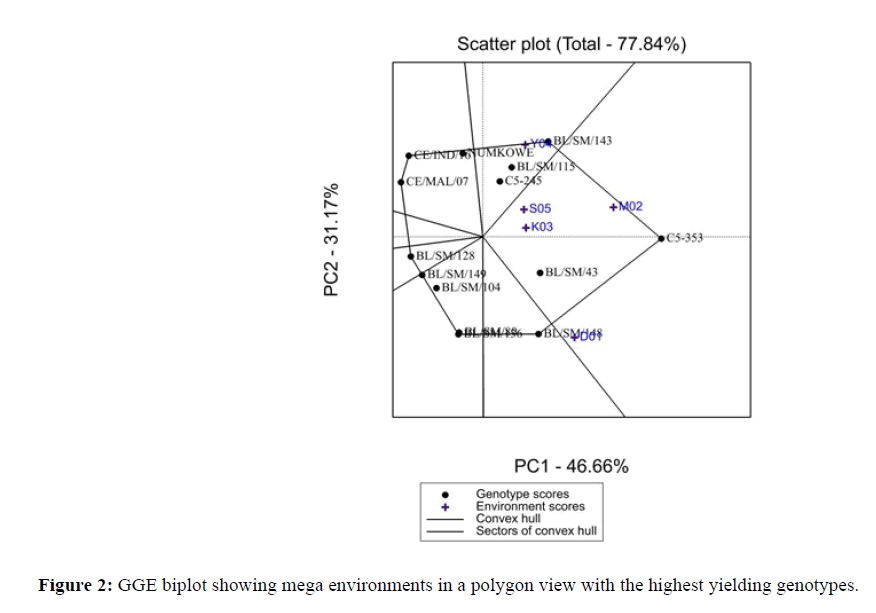
Figure 2: GGE biplot showing mega environments in a polygon view with the highest yielding genotypes.
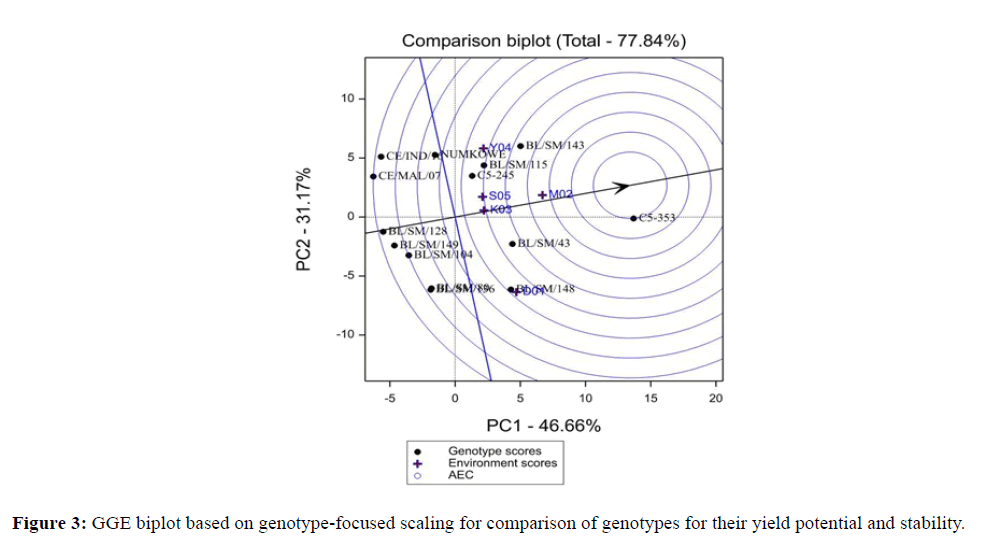
Figure 3: GGE biplot based on genotype-focused scaling for comparison of genotypes for their yield potential and stability.
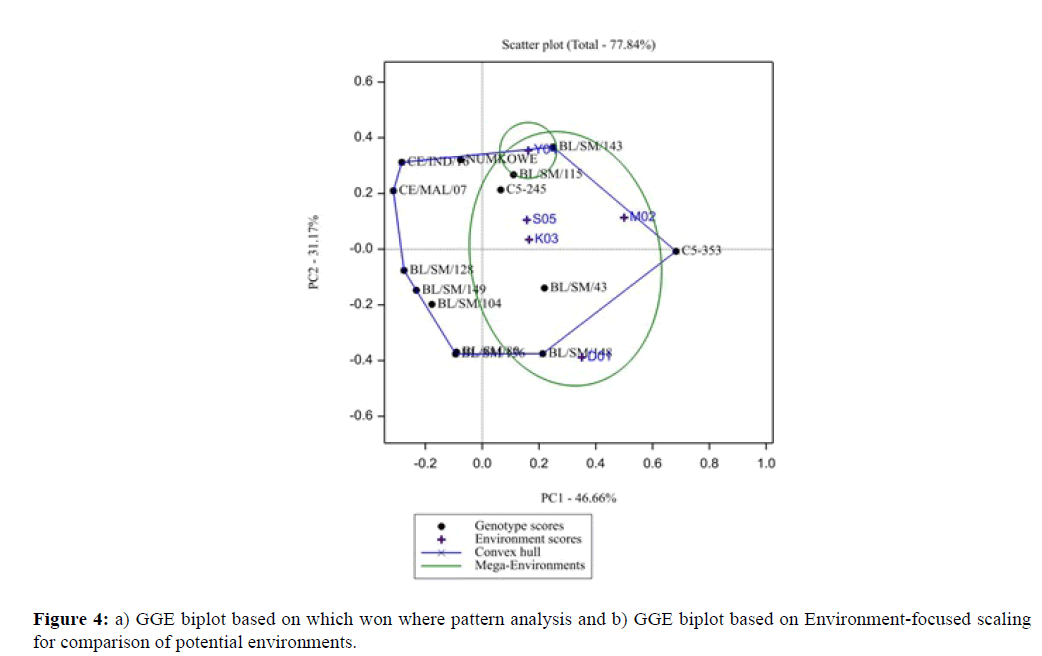
Figure 4: a) GGE biplot based on which won where pattern analysis and b) GGE biplot based on Environment-focused scaling for comparison of potential environments.
Mean Performance and stability of the 13 taro genotypes
Through the comparison analysis in biplot, ideal genotype can be identified which had the highest mean performance and stable in performance across all environments (Figure 3, Tables 3 and 4). The ideal genotype has the highest average value of all genotypes and is stable with low PCA scores (PCA scores near zero) (Table 3). An ideal genotype is defined by having the greatest vector length of the high yielding genotypes with PCA near zero GEI, as represented by an arrow pointing to it (Figure 3). Additionally, it does not exhibit any cultivar by environment interactions and genotype by environment interactions hence broad adaptations [23,24] and can be used as a reference for genotype evaluation [25]. In view of this, PNG advance breeding line C5-353 was the most ideal genotype (high mean yield) and small absolute PC2 scores (-1.6) (high stability). It could also have the greatest commercial success because it showed highest stability. Other desirable genotypes such as BL/SM/143, 148, 43 and C5-245, (Figure 3 and Tables 3 and 4) could be included however, 115 could be included for further evaluation for further selection. A genotype is more desirable if it is located closer to the ideal genotype. Thus, using the ideal variety as the centre, concentric circles were drawn to help visualize the distance between each genotype and the ideal genotype type. Because the units of both PC1 and PC2 for the genotypes are the original unit of yield in the genotype-focused scaling (Figure 4), the units of the average environment Coordinate (AEC) abscissa (mean yield) and ordinate (stability) should also be the original unit of yield. The unit of the distance between genotypes and the ideal genotype, in turn, is the original unit of yield as well.
Discriminating ability and Representativeness of the test environments
One of the fundamental and intrinsic features of a GGE biplot analysis is its ability to show the which-won-where pattern of a genotype by environment (Figure 5). In the GGE biplot, it grouped with the higher yielding environments. In the “which one wins where’’ scenario the five environments used in this evaluation trial formed two mega-environments with different winning genotypes. This tendency was identified using the scatter plot with polygon bisect and confidence ellipse (Figure 5). Genotypes C5-353, BL/SM/148 and 143 were winning genotypes for environments (D01, M02 and Y04) while the BL/SM/115,143 and C5-245 were the wining genotypes for environments (S05, Y04 and K03) which was similar to AMMI analysis output respectively.
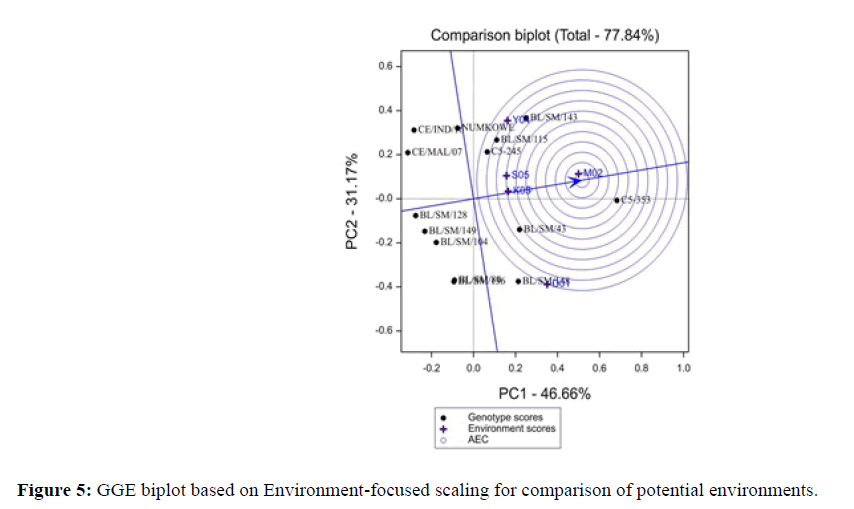
Figure 5: GGE biplot based on Environment-focused scaling for comparison of potential environments.
The PC1 and PC2 were also useful in obtaining ideal environments. The Such environments are always characterised by large PC1 scores, which is very in discriminating genotypes in terms of genotype main effects with small contribution from PC2 scores (more representative of the overall environments) [26-28]. The ideal location should also have the capacity to differentiate the genotypes and representative of the target location. In this study, representativeness referred to as average location, which was used as a reference or benchmark. M02 was the most ideal environment as identified from the GGEbiplot (Figure 6). The AMMI analysis also identified the best environment as M02, which had the highest PCA1 and the best IPCA2 values of 0.01 and -0.14 respectively (Table 6). However, high PCA show unstable yields, which could be used in the selection, site for genotypes to be grown in specific environments [24]. Thus, this study identified the locations, which optimize genotype selection in the basis of their discriminating ability and representativeness.
| Genotype | Gmean | Gm rank | ASV | YSI | IPCA1 | IPCA2 |
|---|---|---|---|---|---|---|
| BL/SM/104 | 8 | 10 | 1.25 | 21 | -0.59 | 0.85 |
| BL/SM/115 | 11.1 | 4 | 1.98 | 14 | 0.94 | 0.8 |
| BL/SM/128 | 7.6 | 12 | 0 | 26 | 0.01 | -0.03 |
| BL/SM/136 | 8.8 | 9 | 3.54 | 13 | -1.35 | 0.87 |
| BL/SM/143 | 12.6 | 2 | 3.72 | 5 | 1.19 | 1.25 |
| BL/SM/148 | 11.2 | 3 | 4.77 | 5 | -1.69 | -0.67 |
| BL/SM/149 | 7.1 | 14 | 0.39 | 27 | -0.37 | -0.43 |
| BL/SM/43 | 10.9 | 6 | 1.18 | 18 | -0.82 | 0.41 |
| BL/SM/80 | 8.9 | 8 | 2.88 | 15 | -1.32 | 0.49 |
| C5-245 | 11 | 5 | 2.82 | 13 | 0.79 | -1.37 |
| C5-353 | 14.9 | 1 | 3.45 | 6 | -0.8 | -1.58 |
| CE/IND/16 | 7.7 | 11 | 4.98 | 12 | 1.51 | -1.23 |
| CE/M AL/07 | 7.5 | 13 | 2.04 | 22 | 1.16 | 0.03 |
| NUMKOWE | 9.5 | 7 | 3.04 | 13 | 1.33 | 0.58 |
Table 6: Cultivaror genotype mean (corm yield (t/ha) , first and second IPCA, AMMI stability values (ASV),Yield stability index (YSI) and ranking orders of the 13 genotypes including the check evaluated.
The result of this study showed that taro corm yield performance was greatly influenced by the environment while GEI contributed least phenotypic variation. Further testing of these taro genotypes in more seasons across the country could enhance breeding efficiency with reference to genotype stability and adaptation across environments. The Genotype C5-353 was ideal followed by BL/SM/148, 43, 143 and C5-245. This genotype should be recommended for release as they have already shown their wide adaptability and yield stability across environments and farmer acceptability. In addition, further testing and studies on food properties of these genotypes would enhance their release given the need for increase taro production, productivity, and diversity to enhance resilience to food security in Papua New Guinea.