Research Article - Journal of Natural Product and Plant Resources ( 2017) Volume 7, Issue 2
The evolution of phenolic content, polyphenoloxidase (PPO) and peroxidase (POD) activity in cotton leaves treated with ethephon and MeJA was studied. Pyrocatechol and guaiacol were used as a substrate for the determination of PPO and POD activity, respectively. The total phenol content was compared under the action of both oxidation enzymes in cotton leaves. The results showed that phenolic content and PPO activity increases with of elicitor’s application on the cotton leaves. Thus, compared to the control, it is higher in ethephon-treated plants followed by MeJA-treated plants. Co-treatment of leaves with these both elicitors stimulates more phenolic compounds production as well as PPO activity thus showing a synergistic or cooperative effect. However, with respect to POD activity, elicitors have an inverse effect compared to PPOs. This indicates that catecholase activity overlapping cresolase activity under the action of MeJA or ethephon in cotton. POD is mobilized to induce the biosynthesis of lignin that constitutes a physical barrier against the pathogens. Furthermore, a part of PPO evolves in a similar way as the total phenol content would indicate an abundance of substrate of this enzyme in treated-plants with elicitors. The phenols synthesized under the action of MeJA and of ethylene could be polyphenolic nature. Concomitant treatment of plant with MeJA and ethylene in the form of ethephon is better than that of MeJA followed by ethephon
Cotton, Leaf, Phenolic compounds, Polyphenoloxidase, Peroxidase.
Cotton is a tropical plant with four currently cultivated species. Gossypium hirsutum is the main Cotton species that is actually cultivated in the world [1]. This plant is cultivated for its fibers which constitute the main raw material in textile industry. Cotton seeds are also used in human and animal nutrition [2]. With 80%, USA, China, India, Uzbekistan and Pakistan are the greatest producers over the world production. African countries account with 8% of this production and the Ivory Coast with 400 000 tons of cotton seeds is the fifth African producer in the South of Sahara after Burkina Faso, Mali, Benin and Cameroun [3,4]. Cotton is an important source of financial resource. In Côte d'Ivoire, it represents the fourth product of exportation after cocoa, rubber and cashew nut [5]. However, the cotton seeds production is affected by the damage of pests and various diseases [6]. In West Africa, including Côte d'Ivoire, cotton diseases constitute the main cause of production loss estimated between 15 at 25%. During unfavorable years, a poorly controlled or uncontrolled parasitism can cause more than 50% production losses and sometimes, this can lead in total destruction of production potential [7]. In tropical climate, chemical control is the main control strategy to fight the impact of parasitism on cotton growing. Unfortunately, pesticides are increasingly indexed to toxicity issue, environmental pollution, health and even destruction of biodiversity [8]. In this context, it becomes necessary to find more efficient ways for the development of sustainable agriculture. One of these solutions consist in allowing plants to self-defend or to stimulate own defenses rather than fighting directly the aggressor [9,10].
Many studies showed that application of elicitor on plant activate its defense response and also leads to increased resistance to pathogens [11,12]. Elicitors are molecules secreted by microorganisms, derivatives from fungal cell wall of bacteria and/or host plant [13]. The eliciting properties could be attributed to some analogous compounds or derivatives of natural molecules such as salicylic acid, ethylene and methyl jasmonate [8,14]. In addition, studies showed that methyl jasmonate and ethylene could synergistically activate some defense genes, leading to an increase resistance face to pathogenic attack [15,16]. According to Ahuja et al. [12] and Belhadj et al. [17] phenolic compounds accumulate in adjacent tissues of necrotic areas. This suggests that phenolic compounds could play a role in plant defense [18]. Cotton produces a large number of these compounds which are critical in plant disease resistance [10]. The works of Konan et al. [10] and Zhu et al. [19] showed an increase in the rate of phenolic compounds production in cotton plants during the interaction of cotton-Fusarium. Vessere [20] reported that degradation or oxidation of phenols has a bactericidal action while Nicholson andHammershmidt [21] observed antifungal action of this product. In plants, peroxidase and polyphenoloxidase are the main enzymes involved in oxidation reactions of phenolic compounds [22].
This work was carried out to study the effect of methyl jasmonate (MeJA) and ethylene (under ethephon form) as well as the interaction (MeJA + ethephon) on the activity levels of PPO and POD and total phenolic compounds in cotton leaves.
Plant material
Plant material consisted of cotton cultivar Y331B seeds was provided from Ivorian Company for Textile Development (ICTD). This cultivar has a high sensitivity to Fusarium wilt.
Chemicals
Dowex-2, ethanol, ethephon, Folin-Ciocalteu’s reagent, gallic acid, glycerol, guaiacol, hydrogen peroxide, mercaptoethanol, methanol, methyl jasmonate, pyrocatechol, polyethylene glycol 6000 (PEG 6000), sodium carbonate, sodium hypochlorite, sodium phosphate, sodium thiosulfate, sulfuric acid and Triton X-100 were purchased from Sigma Co. (Sigma Aldrich, Germany). All the solvents used were of analytical grade.
Methods
Obtaining cotton plants
Cotton seeds were delinted with concentrated sulfuric acid. Plump and mature seeds were sterilized by rapid dipping (1 min) in ethanol (70%) followed by immersion for 20 min in sodium hypochlorite (3.6%). Seeds were sown in pots containing sterilized soil by autoclaving at 121°C for 30 min under 1 bar pressure. They were in vivo germinated in a shelter covered with transparent plastic film for two months.
Preparation and application of elicitors
Methyl jasmonate (MeJA) and ethephon were previously dissolved in 1.0% of ethanol containing 0.1% Triton X-100 as a surfactant. MeJA and ethephon were prepared aseptically in a laminar flow hood and solvents used sterilized by filtration (0.22 μm). The leaves of each seedling were sprayed with 10.0 mL of elicitor solution. The control lot was treated with solution containing 1.0% ethanol and 0.1% Triton X-100 without elicitor. Watering of the seedlings was ensured according to the moisture of the substrate. Each treatment consisted in three replicates. One hundred and fifty seedlings of cotton were used in this study. Seedlings were sprayed with a 5.0 mM of MeJA and 5.0 g/L of ethephon [23]. A co-treatment with MeJA and ethephon was also achieved. Ten seedlings were used for each type of treatment. After 72 h of incubation, leaves of seedlings were harvested and lyophilized.
Phenolic compounds
One hundred (100) mg of freeze-dried leaf were added to 10 mL of methanol and incubated overnight at 4°C in a blender. Sample was centrifuged at 10,000 rpm for 10 min. Supernatant was collected and used as phenolic extract to be analyzed.
Determination of total phenolic content
The total phenolic content of extract was determined using Folin-Ciocalteu’s reagent according to the method of Siriwoharn et al. [24]. Briefly, 500 μL of crude extract were made up to 3 mL with distilled water, mixed thoroughly with 0.5 mL Folin-Ciocalteu reagent for 5 min, followed by the addition of 2 mL sodium carbonate 20% (w/v). The mixture was allowed to stand for a further 30 min in darkness, and absorbance was measured at 765 nm. The total phenolic content was calculated from the calibration curve, and the results were expressed as mg of gallic acid equivalent per gram of dry weight. All measures were performed in triplicate.
Peroxidase and polyphenoloxidase extracts preparation
Enzyme extractions were performed following the method described by Kouakou et al. [25]. Approximately, 0.5 g of freeze-dried leaves were ground in a porcelain mortar in the presence of in ice-cold sodium phosphate buffer (0.1 M ; pH 7.0), with 1% triton X-100, 0.5% PEG 6000, 0.25% sodium thiosulfate, 15% glycerol and 15 mM mercaptoethanol. The resulting suspension in buffer was centrifuged at 10,000 rpm for 20 minutes at 4°C. The supernatant designated as soluble enzymatic extract was collected and stored at -25°C in freezer. To get rid of inhibitor ions, Dowex-2 was added to the crude enzymatic extract and the mixture was incubated during 30 min at 4°C. The supernatant was collected and centrifugation carried out under the same prior conditions. Then, the supernatant obtained contains the purified enzymatic fraction and was immediately used to determine the activities of the following enzymes.
Determination of peroxidase activity
The activity of peroxidase (POD; EC 1.11.1.7) was assayed by the method of Kumar and Khan [26] with some modifications. The reaction mixture used for estimating the POD activity contained 1.8 mL sodium phosphate 0.1 M (pH 7.0), 0.5 mL guaiacol 0.01 M, 0.5 mL hydrogen peroxide (H2O2) 0.01 M and 0.2 mL of the enzyme extract. This mixture was incubated for 10 min in darkness at 25˚C.Then, the reaction was stopped by adding 1 mL sulfuric acid (H2SO4) 2.5 N. Colour development was measured by UV spectrophotometer at 470 nm as a change in absorbance of 0.001/min against a reagent blank prepared by adding the extract after the addition of 2.5 N H2SO4 at the zero time (before reaction beginning). POD activity was expressed in mmol per gram of dry weight (mmol/g/dw) by considering that the molar extinction coefficient of the formed product is 26.6 cm-1 [27]. POD activity was assayed in triplicate.
Determination of polyphenoloxidase activity
Polyphenoloxidase (PPO; EC 1.14.18.1) activity quantification was done following the method described by Murata et al. [28] with some modifications. The reaction mixture used for estimating the PPO activity contained 2.4 mL sodium phosphate 0.1 M (pH 7.0), 0.4 mL pyrocatechol 0.13 M and 0.2 mL of the enzyme extract. The mixture was incubated for 10 min at 25˚C. PPO activity was determined by measuring the absorbance at 420 nm against a reagent blank containing only 3 mL of reaction mixture without substrate (pyrocatechol). An absorbance decrease of 0.01 per minute was considered 1 unit of PPO activity [29]. The linear portion of the absorbance vs time curve was used to determine the initial rates [25]. PPO activity was expressed in mkat per gram of dry weight (mmol substrate converted/s/g dw). PPO activity was assayed in triplicate.
Statistical analysis
The experiment design was as a completely randomized one with five replicates (each replication including 10 plants). ANOVA was performed for the experiment using Statistica software (release 7.5). Means of data were compared by Duncan’s Multiple Range Test. Differences at P ≤ 0.05 were considered significant. For percentages, which are nonparametric values, Kruskal-Wallis test was used at P ≤ 0.05.
The results show that phenol content of cotton leaves is significantly greater in MeJA-treated plants than in those with ethephon (Figure 1). These results are in agreement with those of [30] who showed that MeJA strongly stimulates the accumulation of phenolic compounds unlike ethephon in the vine.
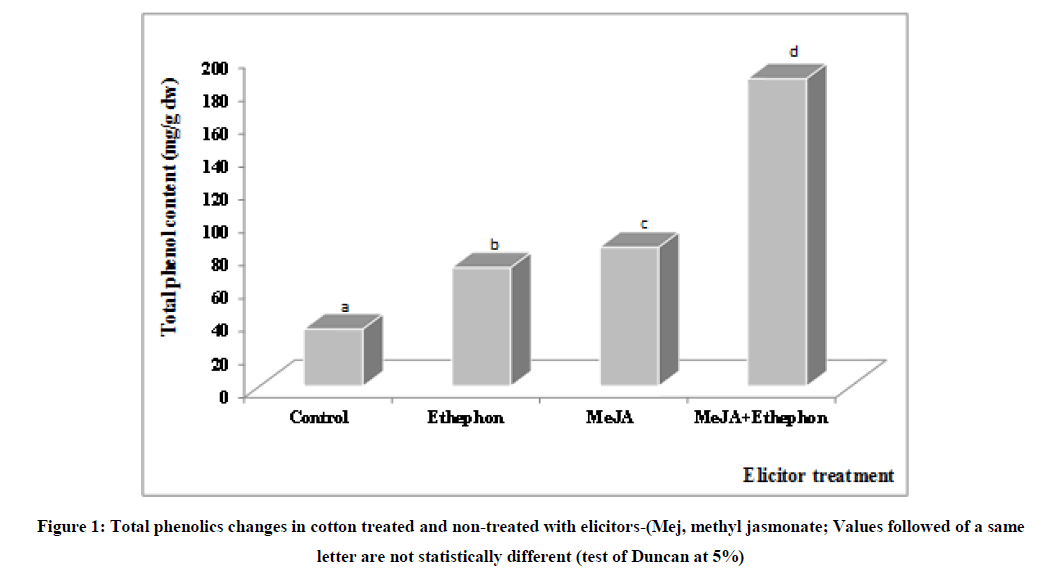
Figure 1: Total phenolics changes in cotton treated and non-treated with elicitors-(Mej, methyl jasmonate; Values followed of a same letter are not statistically different (test of Duncan at 5%).
In addition, Zhao et al. [31] reported that the accumulation of phenolic compounds in plant activates defense reactions; what provokes resistance to pathogen attacks. Thus, MeJA would activate the cotton plants defense reactions more than ethephon (ethylene). Furthermore, Diaz et al. [32] showed that ethephon stimulates the natural defenses of tomato plants by inducing resistance against Botrytis cinerea. These results seem to suggest a varietal dependence in action of MeJA and ethephon. In our study, ethephon seems to less activate the natural defenses of cotton than MeJA as already indicated by Faurie et al. [8] in the vine.
Moreover, leave co-treatment with MeJA and ethephon allowed an exponential increase in the phenolic content compared to single application of MeJA. So, the concomitant application of MeJA and ethephon has a synergistic effect on the accumulation of phenolic compounds in cotton. This cooperative effect of MeJA and ethephon on the biosynthesis of phenolic compounds was already reported in many plants [8,10,33]. Both elicitors (MeJA and ethephon) seem to be inserted in cascade reactions which lead to the mobilization of phenolic compounds having a phytoalexin nature. These both elicitors would trigger larger defense mechanisms when used together than individually. These results suit to those of [15] who showed that MeJA and ethylene can sinergically act on some genes coding for phenolic biosynthesis.
Phenolic compounds are known for their antifungal properties and their involvement in plant resistance to pathogens was already reported [12,34]. It is reported that phenolic compounds are accumulated in tissues adjacent to necrotic areas, suggesting that these compounds may be defensive ones [10,14,18,35]. Besides, Vessere [20] noted that phenolic product has a bactericidal action, while Nicholson and Hammerschidt [21] reported their antifungal action. These observations suggest that oxidation enzymes of phenolic compounds play an important role in plant defense [36,37]. If the activities of biosynthesis enzymes play a regulating role in the kinetics of phenolic accumulation, they are not however the only ones. Indeed, the phenol contents measured at a given stage are the result of a balance between biosynthesis and especially oxidative degradation [38].
In plants, Tognolli et al. [39] showed that peroxidase (POD) and polyphenoloxidase (PPO) are involved in these oxidations. Analysis of phenolic compound oxidation enzyme activity shows a decrease activity of POD and an increase activity of PPO in elicitor-treated plants compared to control (Figure 2). The POD activities are inversely related to the phenol content whereas those of PPO evolve similarly as the phenol content in the treated-plants. The results show that there is more substrate for PPO than POD.
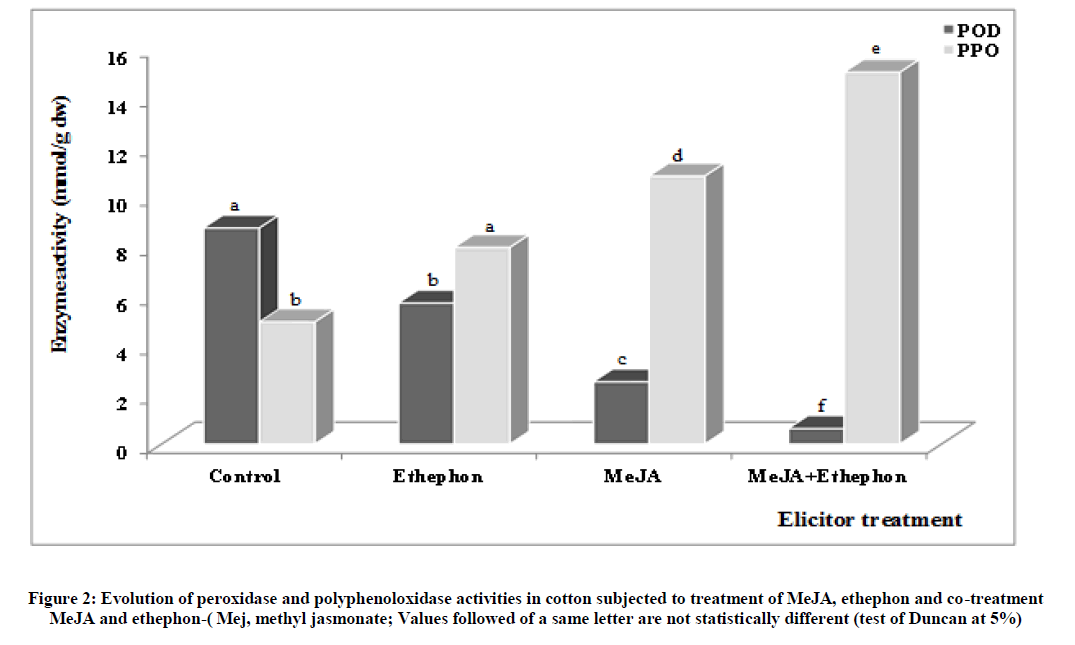
Figure 2: Evolution of peroxidase and polyphenoloxidase activities in cotton subjected to treatment of MeJA, ethephon and co-treatment MeJA and ethephon-( Mej, methyl jasmonate; Values followed of a same letter are not statistically different (test of Duncan at 5%).
This seems to justify the high activity of PPO. Konan et al. [10] reported that MeJA and ethephon stimulate the phenolic biosynthesis in cotton. Thus, this biosynthesis could be more oriented towards the polyphenols formation than that of monophenols under the effect of elicitors. These results are in agreement with those of Constabel et al. [40] who revealed an increase activity of PPO activity in presence of elicitor. Our results showed that PPO activity is higher than that of POD except at the control level (Table 1). This important activity of POD is justified by a low availability of phytoalexins under phenolic form to defend plant against pathogens attack.
| Contribution to phenols oxidation (%) | |||
|---|---|---|---|
| Treatment | Ratio PPO/POD | POD | PPO |
| Control | 0.56 ± 0.002a | 63.88 ± 0.50e | 36.12 ± 0.80f |
| Ethephon | 01.39 ± 0.05b | 41.74 ± 0.70f | 58.25 ± 0.60e |
| MeJA | 04.31 ± 0.03c | 18.83 ± 0.30g | 81.17 ± 0.90i |
| MeJA + Ethephon | 24.20 ± 0.10d | 03.97 ± 0.20h | 96.03 ± 0.50j |
Table 1: Ratio of polyphenoloxidase and peroxidase activities and contribution rate of each enzyme to phenols oxidation
The preformed phenols of cotton would oxidize to equip plants with quinones those antifungal or antibacterial actions has already been reported by several authors [21,41,42]. Analysis of PPO/POD ratio indicates that the PO activity is 24.20 times greater than that of POD in co-treated plant with MeJA and ethephon, followed by MeJA (4.31) and ethephon (1.39).
This result reveals an important activity of PPO compared to that of POD. Indeed, the rate of PPO contribution to phenol oxidation, which was 36.12% in control, increased to 58.25% with ethephon and then to 81.17% with MeJA to reach 96.03% with co-treatment. The oxidation of phenols by PPO leads to the formation of diquinones whose beneficial effect in the plants defense was demonstrated by several authors [42,43]. This intense activity of the PPO in elicitor-treated plants suggests a strong increase of the polyphenols in these plants.
POD, peroxidase; PPO, polyphenoloxidase; MeJA, methyl jasmonate; for contribution to phenol oxidation, the values followed of a same letter are not statistically different (test of Kruskal-Wallis at 5%); for PPO and POD ratio, the values followed of a same letter are not statistically different (test of Duncan at 5%).
Indeed, known under diverse names such as tyrosinase, phenolase, catechol oxidase, catecholase, monophenoloxidase, o-diphenol oxidase or o-phenolase, depending on substrate specificity, PPO has monophenolic and diphenolic activities [44]. Monophenolic (cresolase) activity is a hydroxylation of monophenols to o-diphenols, while that of diphenolic (catecholase activity) is an oxidation of o-diphenols to diquinones [41,45]. There would therefore be a good availability of diphenols; what seems to justify the higher activity of PPO observed in cotton leaves [46]. The decrease activity of POD in the treated plants is in agreement with that reported by Ziouiti et al. [47]. Furthermore, these authors showed that the increase activities in POD coincided with a decrease in phenol content and conversely in the date-tree root infested with pathogens. However, Dai et al. [48] reported high POD activity at the cell walls and intercellular spaces in cotton leaves infected with Xanthomonas campestris. According to Huckelhoven [49], this increase activity of POD is due to their participation in antimicrobial hydroxyl radicals' production. In our study, decreased POD activity in treated plants may indicate a low participation of POD in the removal of hydrogen peroxide (H2O2) [50,51]. Indeed, the main enzymes in the detoxification of H2O2 are catalase and POD [52]. Catalase detoxifies most of H2O2 produced while POD neutralizes the H2O2 molecules undestroyed by catalase. However, according to Zhang and Kirkham [53], the role of POD in plant protection against H2O2 is minor. This suggests that POD could have another physiological function. Besides, PODs have a multifunctional role [54]. They participate in a wide range of physiological processes such as auxin metabolism, lignin biosynthesis and suberin formation [55-59]. POD catalyzes the oxidation of monophenols to monoquinones which are fungitoxic [21]. Furthermore, EL Bellaj and EL Hadrami [42] reported that POD activity in resistant plants increases during the attack of pathogens in order to combat their invasion. In our study, decreasing activity suggests that elicitors do not induce stress in cotton as reported by many studies [38,56-60]. Elicitors would attach to a plant cell receptor, triggering a cascade of actions that results in the synthesis of defense compounds such as phenolic compounds [11]. So, using an elicitor may reduce the amount of pesticides needed to protect a crop. This technique of phytosanitary control is called "stimulation of the natural defenses of plant".
Activity of POD also decreases according to the treatment with elicitor. However, unlike PPO, the participation of POD in phenolic oxidation in treated plants is low. This supposes a weak monophenolic content in these elicited plants compared to control. So, untreated-plants would possess more monophenolic compounds with regard to elicited plants, hence more POD activities. However, POD would more participate in the preformed defense of plants such as lignin or leaf cuticle formation than in induced defense. That could be the case for PPO in this study. This, with the aim of strengthening the cell walls to create a structural barrier against a possible penetration of pathogens in cells to infest the plant [54,61,62]. In leaves, lignin content is low because it can be estimated between 3 and 5% [63,64]. This situation may explain the low POD activity observed in the elicited-leaves. It is worth noting that the high activity of PPO suggests that these enzymes would have cresolase activity in cotton. However, this activity is transformed into catecholase one under the action of elicitors. The elicitation would cause a biosynthesis of polyphenols. The plants co-treated with MeJA and ethephon increased PPO activity while it decreased the POD one.
This stimulation of cotton defenses seems to be based on the same principle as vaccination. Indeed, the cotton natural defenses would be activated before contamination by pathogens. So, the cotton plants should be put in contact preventively with MeJA and ethylene which are able to activate their natural defenses by synthesizing defensive compounds. Some of these compounds would oxidize to be more toxic to pathogens [41,44,65]. So, it would allow the plant to be in a resistance state against a pathogen to which it would normally be susceptible (phyto-vaccination). Plants defense impact is further enhanced when they are treated concomitantly with MeJA and ethephon. Showing a synergistic effect of this co-treatment on the phenolic compounds production. It has already been reported that MeJA is a molecule that elicits the biosynthesis of phenolic phytoalexins [8,10,62,66] against pests such as Fusarium oxysporum f. sp. vasinfectum, causal agent of Fusarium wilt in association with ethephon.
Cotton plants treatment single or simultaneously with MeJA and ethephon, increase PPO activity while they decrease POD activity and evolves in the same way as phenol content compared to control plants (untreated). This indicates a strong catecholase activity in cotton to the detriment of cresolase activity. So MeJA and ethephon can be considered as elicitors of cotton defense. However cotton co-treated with the both elicitors accentuates the decrease of POD activity and increase of PPO activity more than MeJA or ethephon used alone. Thus, polyphenols are more mobilized when treating plants with elicitors. The high phenols content observed in treated-plants by elicitors show that plants are equipped with defense compounds. So, PPO activities are oriented towards the phenols oxidation to give toxic compounds. As for the POD, their oxidation activity is mimed in favor of their lignin biosynthesis activity in order to create a structural barrier against pests. Finally, it should be noted that the co-treatment with the two elicitors is better followed by the MeJA and the ethephon in comparison with the control which is untreated-plant.