Research Article - Journal of Natural Product and Plant Resources ( 2017) Volume 7, Issue 1
In Algeria, Aleppo pine is a major forest tree by its surface and its importance in reforestation programs. Despite this, the species is not fully known especially in its genetic constitution and characteristics. In this study, 49 open pollinated families from the Aleppo pine stands in the area of Algiers were studied. Morphological and adaptive traits were investigated on one and two-year old seedlings. Results showed a relatively low differentiation among stands and, conversely, a significant variation between families within sands. Implications for the selection are discussed on account of climate changes.
Aleppo pine, stand, family, seedlings, adaptive, morphological traits.
Aleppo pine (Pinus halepensis Miller) is a widespread-circum-mediterranean species whose range spanned 28 degrees of longitude and 15 degrees of latitude. However, this species is mostly found in western Mediterranean [1-5], where it constitutes a common element of the landscapes; in eastern Mediterranean, its populations often are scattered and relic [6,7].
In Algeria, this important forest tree covers about 850,000 ha [8,9] in coastal and inland areas (Figure 1), where populations are subjected to various selection pressures and, for centuries, were shaped by man and fire. Fire is an important factor favouring the spread of the species. In spite of its importance as a major species in planting programs, Aleppo pine, in Algeria, is poorly known, with regard to its geographic and/or genetic variability. Little was made to investigate these important features for the use and the conservation of the species. Nevertheless, a few studies suggested the existence of a geographic and ecotypic differentiation among populations [9]; however, these rare studies dealt with populations in situ and, thus, did not allow determine whether the observed differences were of genetic or environmental nature. Furthermore, not any of these studies dealt with individual and/or within population variation, hence keeping a wide gap in our knowledge about genetics and population structure of this species. We attempted, since a few years, to fulfil, in part, such a lack of data in establishing statistical experiments and designs to investigate among and within-population variability of the species. Morphological and growth traits are tightly related to the fitness of individuals and populations; thus, and within current climate warming, a good knowledge of how they adapt themselves to changes are of key importance to maintain forests, particularly in arid and semi-arid zones. Hence, testing individuals and populations in common environments and well-defined experimental designs represents a challenge for the future.
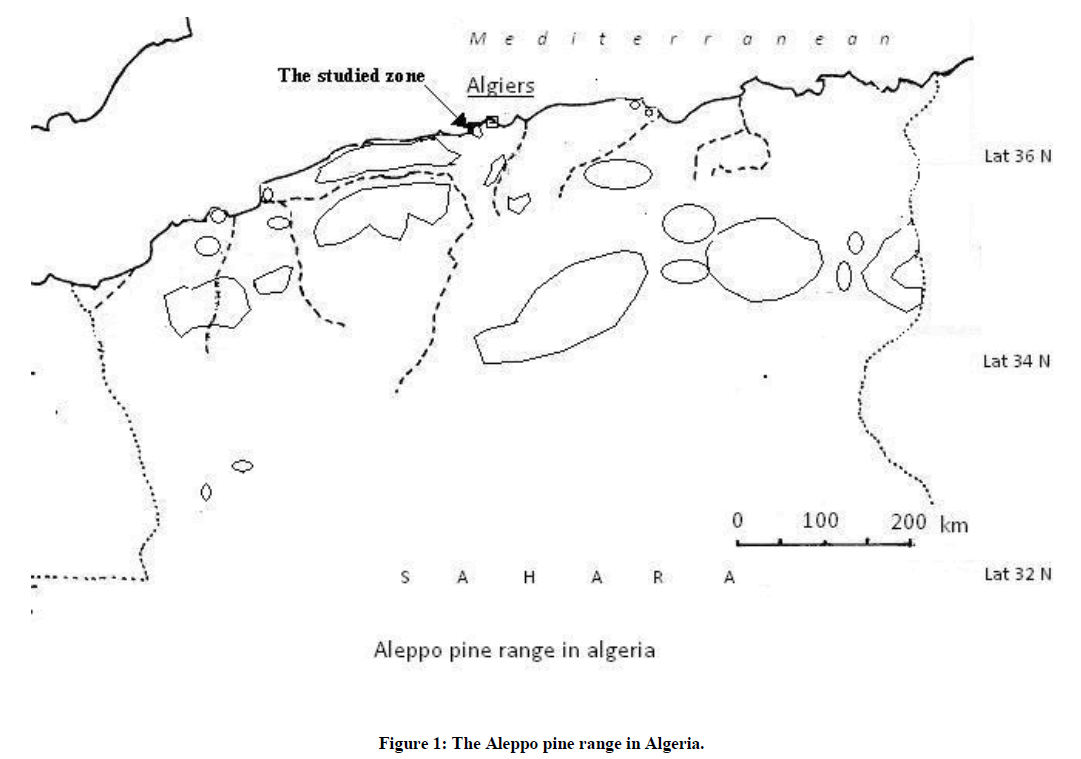
Figure 1: The Aleppo pine range in Algeria.
Towards these objectives, we collected 22 provenances countrywide distributed and 49 open pollinated families from the Aleppo pine stands westward of Algiers. Morphological and adaptive traits, as well as a few isozyme loci, were investigated. Papers were published on cone, seed and Plantlet traits [10], and isozymes [11]. The present work deals with seedling morphological and growth attributes at ages 1 and 2 on 49 open pollinated families from stands westward of Algiers.
Forty nine (49) open pollinated families of 8 stands of Aleppo pine from the western coastal region of Algiers were included in the study (Figure 2). This region is particularly interesting to study because of its ecological diversity and the occurrence of a typical "Mediterranean coastal facies" of the species, first described by Maire [12]. The prospected zone extends over about 15 km parallel to the Mediterranean coast and over about 3 km in depth in the back country. Geographic and ecological data as well as the number of families for each stand are presented in Table 1. The distance between collected trees was 80-100 m in order to minimize collection on related individuals. Individuals retained as mother trees were not selected in order to provide more representative samples.
| Stand | Number of collected families | Number of seedlings | Soil | Order of size | Long* (km) | Alt.** (m) |
|---|---|---|---|---|---|---|
| Subcoastal Douaouda (Doua1) | 4 | 80 | Deep red silty clayey | A few hundreds of trees | 0 | 46 |
| Coastal Douaouda (Doua2) | 1 | 20 | Deep sandy (Dune) | A few dozens of trees | 1 | 10 |
| Mazafran coastal (Mazafran1) | 3 | 60 | Deep red silty clayey | A few scattered trees (A dozen) | 2.5 | 17 |
| Mazafran subcoastal (Mazafran2) | 6 | 119 | Deep red silty clayey | Thousands of trees | 3.5 | 17 |
| Zeralda subcoastal (Zeralda1) | 7 | 140 | Deep brown, clayey | Thousands of trees | 4.5 | 54 |
| Zeralda coastal (Zeralda2) | 6 | 120 | Deep sandy (Dune) | Thousands of trees | 7.5 | 9 |
| Zeralda-game reserve (Zeralda3) | 14 | 280 | Deep red silty clayey | Thousands of trees | 10.5 | 20 |
| Bouchaoui | 8 | 158 | silty clayey | Thousands of trees | 15 | 110 |
| Total | 49 | 935 |
*Long. = kilometric longitude; **Alt. = average altitude of the collected trees.
Table 1: Characteristics of stands and sample sizes.
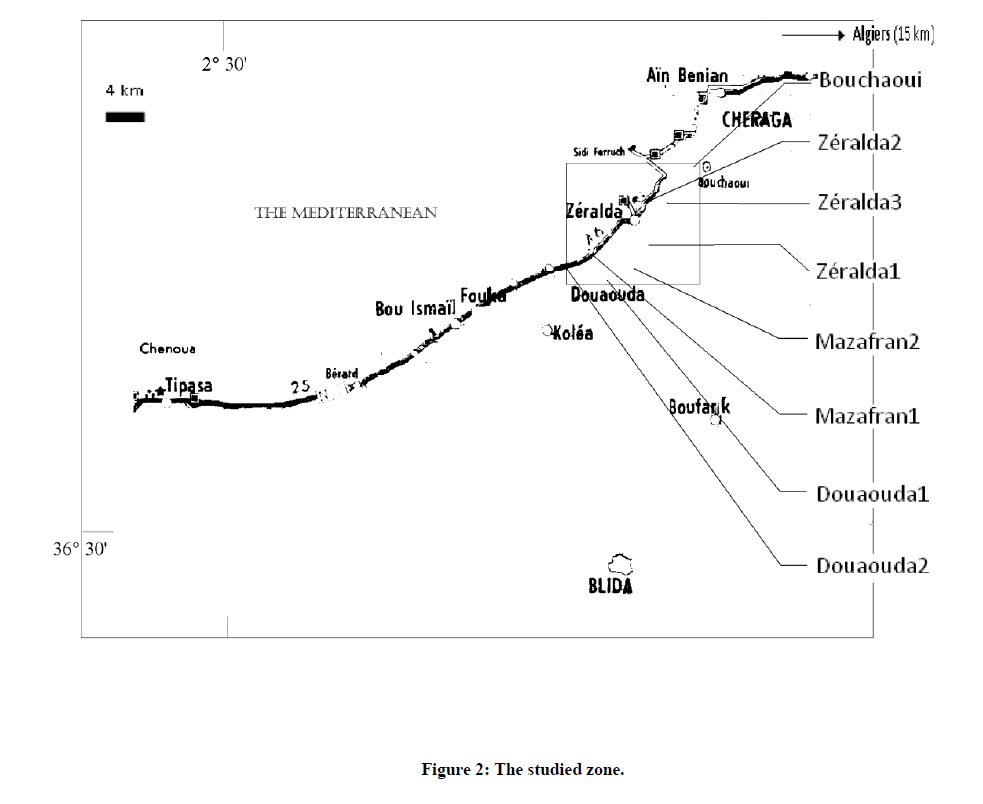
Figure 2: The studied zone.
Globally, the climate of the zone is sub-humid with, however, a gradient of annual rainfall from western (628 mm) to eastern (717 mm) boundaries. A relative variety of soils is encountered within this region, but deep sandy and red silty or clayey soils are the most widespread ones.
Seeds were sown in nursery in bottomless polythene bags (9 cm diameter, 20 cm height) at the rate of 2 seeds/bag and 20 replicates per family. At the age of 8 months, the seedlings were transplanted in 10-liter pots arranged in a random-complete-block design constituted by 20 blocks, each block containing one potted-seedling per family; a block therefore contains 49 seedlings, randomly drawn.
The traits assessed on seedlings are defined in Table 2. The lines of stomata were observed under a binocular.
| Variable | Stand | F-test | Probability | ||||
|---|---|---|---|---|---|---|---|
| Mean | Min | Max | Stand | Family/ stand | Stand | Family/ stand | |
| TH1 (cm) | 18.21 | 13.04 | 20.45 | 4.06 | 12.93 | 0.002 | <10-6 |
| TH2 (cm) | 34.42 | 30.33 | 36.44 | 2.45 | 3.89 | 0.033 | <10-6 |
| ΔTH (cm) | 15.97 | 14.31 | 17.26 | 0.95 | 1.97 | 0.48 | 0.0003 |
| DB1 (mm) | 3.92 | 3.66 | 4.75 | 2.51 | 2.83 | 0.03 | <10-6 |
| DB2(mm) | 9.47 | 8.74 | 9.99 | 4.3 | 2.6 | 0.001 | <10-6 |
| ΔDB (mm) | 5.56 | 5.03 | 5.81 | 2.46 | 1.4 | 0.033 | 0.052 |
| DB2/TH2 | 0.29 | 0.27 | 0.32 | 3.25 | 3.12 | 0.007 | <10-6 |
| NLSprouts | 10.5 | 9.72 | 11.28 | 0.41 | 7.7 | 0.891 | <10-6 |
| DevVerticality(Degrees) | 3.91 | 1.7 | 5.06 | 1.68 | 1.64 | 0.14 | 0.007 |
| NeedLength (mm) | 8.12 | 7.75 | 8.46 | 1.37 | 1.16 | 0.245 | 0.226 |
| NeedShLength (mm) | 3.1 | 2.89 | 3.77 | 1.18 | 0.9 | 0.333 | 0.636 |
| NeedShLength/NeedLength | 0.39 | 0.37 | 0.44 | 0.59 | 1.05 | 0.758 | 0.379 |
| NStDS | 2.37 | 2.3 | 2.42 | 1.52 | 1.52 | 0.189 | 0.021 |
| NStVS | 6.32 | 5.95 | 6.54 | 0.59 | 1.41 | 0.757 | 0.046 |
| NStM5 | 37.14 | 36.7 | 37.82 | 1.49 | 0.86 | 0.196 | 0.713 |
TH1: total height at age 1; TH2: total height at age 2; ΔTH: height increment (TH2-TH1); DB1: Diameter at the base at age 1; DB2: Diameter at the base at age 2; ΔDB: Base Diameter increment (DB2-DB2); DB2/TH2: slenderness; NLSprouts: number of lateral sprouts; DevVerticality: deviation from the verticality; Need Length: needle length; NeedShLength: needle-sheath length; NStDS : number of lines of stomata on the dorsal side of a needle taken from the upper part of the seedling; NStVS : number of lines of stomata on the ventral side of a needle taken from the upper part of the seedling; NStM5(1): number of stomata on the middle-5cm of the needle taken from the upper part of the seedling.
Table 2: Overall Means, maximum and minimum for stand effect, F-tests and probabilities for stand and family within stand for each trait.
Statistical analyses
Variation in seedling characters was analysed by means of a hierarchical two-way analysis of variance whose model was as follows:
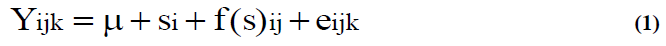
Where, Yijk is the individual (phenotypic) measure, the grand mean, si the (random) ith stand effect (i=1-8), f(s)ij the (random) jth open-pollinated family within the ith stand effect (j=1-49), and eijk the random raw error.
The following parameters were calculated:

Where, 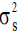 is the stand component of variance,
is the stand component of variance, 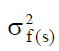 is the open-pollinated family within stand component of
variance and
is the open-pollinated family within stand component of
variance and 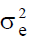 the error variance. Gd measures the extent of between-stand differentiation:
the error variance. Gd measures the extent of between-stand differentiation:

t is the intra-class (family) coefficient from which heritability is derived. Given the nature of the plant material (open-pollinated families) and assuming no significant selfing in Aleppo pine, we took a coefficient of relationship of 1/4. Thus, heritability was calculated as h2 = 4t.
Correlation analyses were made between seedling (age 1 and 2) and plantlet (A few weeks after emergence) attributes with a view to (a very) early selection for growth (from the stage of plantlet) as well as between seedling characters and geographic data for Longitude and Altitude.
A Multivariate Hierarchical Classification was run on family-within-stand means in order to test the degree of conformity between the pre-supposed family membership (to stands) with the clustering obtained by the Hierarchical classification. A coefficient of Conformity (C) between the two structures was calculated as follows:
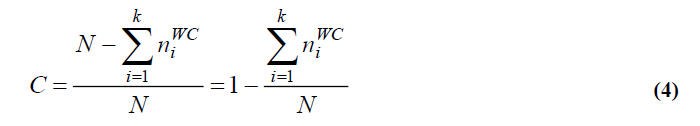
Where, N is the total number of families for all of the stands, k the Number of stands and ni WC the absolute frequency of Wrongly-Classed families in the ith stand. The statistical significance of this coefficient was tested by using a Chisquare test based on a 2 x 2 table opposing the frequencies of the Observed stand-membership of the families and the frequencies of wrongly-classed families (See annex).
The method of Ward was performed to aggregate families into clusters using Euclidian distances.
Geographic differentiation and local variation
Means (For stands) and F-tests (For stand and family-within stand effects) and probabilities are displayed in Table 2. At the stand level, only traits related to growth (Total height and basal diameter at ages 1 and 2, and the basal diameter increment) displayed statistically significant (p<0.05) differences between stands; variability due to stand effect was little if not negligible.
Hence the very low coefficients of differentiation between stands (Gd) which values spanned from 0 to 0.17, i.e. the variance due to stand component explained between 0% to 17% of the total variance depending on the considered trait, suggesting that geographic differentiation was absent or small among stands within the area under study for the group of studied traits. Compared to cone, seed and plantlet traits [10], seedling traits at ages 1 and 2 showed a slightly less geographic differentiation as it is suggested by the mean values of Gd (0.031 for the former vs 0.024 for the latter) (Table 3).
On the other hand, at the family-within-stand level, variation between families was highly significant (p<10-6) for growth traits (Total height, Diameter at the base, Number of lateral sprouts), moderately high (0.001<p<0.05) for form trait (Deviation to verticality) and Stomata traits (Number of lines of stomata on ventral or dorsal sides) and non-significant (p>0.05) for needle traits (Table 2).
Intra-class coefficients, and therefore Heritabilities, were strong for Total height, and the Number of lateral sprouts, which expresses a character of vigor; otherwise, most of traits have very low heritability (Table 3). Such results for vigor might be, at least partially, the expression of maternal effects due to the seed, which can be seen as a common, but transitional, environmental effect accentuating non-genetic resemblance within siblings.
| Traits | Components of Variance | Genetic parameters | ||||
|---|---|---|---|---|---|---|
| δ2s | s2f(s) | s2e | Gd | t | h2 | |
| TH1 (cm) | 5.25 | 9.21 | 15.4 | 0.17 | 0.37 | >1.0 |
| TH2 (cm) | 2.72 | 8.09 | 53.34 | 0.04 | 0.13 | 0.5 |
| ΔTH (cm) | 0 | 3.02 | 59.25 | 0 | 0.05 | 0.2 |
| DB1 (mm) | 0.03 | 0.07 | 0.76 | 0.03 | 0.08 | 0.32 |
| DB2(mm) | 0.11 | 0.12 | 1.4 | 0.07 | 0.07 | 0.29 |
| ΔDB (mm) | 0.03 | 0.03 | 1.66 | 0.02 | 0.02 | 0.08 |
| DB2/TH2 | 0 | 0.0009 | 0.0222 | 0 | 0.04 | 0.16 |
| NLSprouts | 0 | 5.21 | 15.43 | 0 | 0.25 | 1 |
| DevVerticality (degrees) | 0.24 | 0.8 | 24.77 | 0.01 | 0.03 | 0.12 |
| NeedLength (mm) | 0.01 | 0.02 | 2.93 | 0.003 | 0.007 | 0.03 |
| NeedShLength (mm) | 0.003 | 0 | 2.33 | 0.001 | 0 | 0 |
| NeedShLength/NeedLength | 0 | 0 | 0.04 | 0 | 0 | 0 |
| NStDS | 0.008 | 0.03 | 1.134 | 0.007 | 0.03 | 0.1 |
| NStVS | 0 | 0.007 | 0.323 | 0 | 0.02 | 0.08 |
| NStM5 | 0.05 | 0 | 13.1 | 0.004 | 0 | 0 |
| Mean | / | / | / | 0.024 | 0.07 | 0.29 |
δ2s:: Stand variation; 2f(s): Family within Stand variance; 2e: Error variance; Gd: Coefficient of differentiation between stands, t: Intra-class coefficient, h2: Narrow sense heritability. All of effects are considered random. Heritabilities greater than 1.00 can be recorded in some circumstances such as assortative matings (homogamy), G x E correlation and/or G x E interaction. In the case of traits such as height at age 1, non-genetic maternal effects connected to seed (stock) factors could increase the resemblance within progenies, hence overestimating heritability.
Table 3:Variance components and genetic parameters.
Age-age Correlations among characters
Table 4 shows the coefficients of correlation between Seedling (ages 1 and 2) traits at the Stand and the Family-within-Stand levels. As these coefficients are based on means, for stands as well as for families, they are interpreted as genotypic correlations.
| Seedling Traits | Genetic level | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| TH1 (cm) 1 | Stand | 1.00 | ||||||
| Family | 1.00 | |||||||
| TH2 (cm) 2 | Stand | 0.90* | 1.00 | |||||
| Family | 0.76* | 1.00 | ||||||
| ΔTH (cm) 3 | Stand | 0.38 | 0.06 | 1.00 | ||||
| Family | 0.35* | 0.34* | 1.00 | |||||
| DB1 (mm) 4 | Stand | 0.04 | 0.05 | 0.19 | 1.00 | |||
| Family | 0.58*** | 0.2 | 0.55*** | 1.00 | ||||
| DB2 (mm) 5 | Stand | 0.55 | 0.35 | 0.52 | 0.71* | 1.00 | ||
| Family | 0.60*** | 0.48** | 18 | 0.68*** | 1.00 | |||
| ΔDB (cm) 6 | Stand | 0.71* | 0.54 | 0.47 | 0.3 | 0.46 | 1.00 | |
| Family | 0.26 | 0.46** | 0.29* | 0.03 | 0.71*** | 1.00 | ||
| NLSprouts 7 | Stand | 0.38 | 0.21 | 0.43 | 0.65 | 0.59 | 0.02 | 1.00 |
| Family | 0.54*** | 0.12 | 0.58*** | 0.58*** | 0.28 | 0.19 | 1.00 | |
| DevVerticality (Deg.) 8 | Stand | 1.00 | ||||||
| Family | 1.00 | |||||||
| NeedLength (mm) 9 | Stand | -0.07 | 1.00 | |||||
| Family | -0.13 | 1.00 | ||||||
| NeedShLength (mm) 10 | Stand | 0.16 | 0.57 | 1.00 | ||||
| Family | -0.04 | 0.47** | 1.00 | |||||
| NStDS 11 | Stand | 0.27 | 0.01 | -0.54 | 1.00 | |||
| Family | -0.21 | 0.11 | 0.02 | 1.00 | ||||
| NStVS 12 | Stand | -0.21 | 0.44 | 0.45 | 0.33 | 1.00 | ||
| Family | -0.37** | 0.02 | 0.1 | 0.60*** | 1.00 | |||
| NStM5 13 | Stand | -0.64 | -0.44 | -0.14 | 0.24 | 0.11 | 1.00 | 0 |
| Family | -0.26 | -0.1 | 0 | 0.25 | 0.22 | 1.00 | 0 |
Statistically significant coefficients are in bold type (*Significant at 5%; **Significant at 1%; ***Significant at 1‰).
Table 4: Linear Correlation coefficients between Seedling Traits at the Stand (N=8) and the Family-within-Stand levels (N=49) levels.
At the stand level, correlations are globally weak, and the highest coefficients are only significant at a probability of 5%. Positive significant correlations are noticed between traits linked to vigor (Total height at age 1 (TH1) and 2 (TH2), r = 0.90; Diameter at the base of the seedling at age 1 (DB1) and 2 (DB2), r = 0.71). Correlations between the same traits at different stages express a phenomenon of age-age correlation which might be used as criteria for juvenile selection at more or less advanced stages.
The coefficients of correlation between Plantlet and Seedling Traits are displayed in Table 5. At the stand level (N = 8), the only statistically significant coefficient is between LHYP (Length of the Hypocotyl) and NStM5 (Number of stomata on the middle-5-cm of the needle); r(LHYP, NStM5)= -0.71 (p=0.030). Such a negative correlation between the two traits is confirmed at the family within-stand level (N = 49), statistically significant though not very large in value; r= –0.31 (p=0.028). Such a correlation would mean the existence of an antagonistic relationship between the stomata density of the seedling needles at age 2 and the vigor of the plantlet.
| Seedling Traits | Genetic level | Plantlet Traits | |||||
|---|---|---|---|---|---|---|---|
| NCOT | LHYP | LOCOT | SHOCOT | LOCOT/LHYP | COTANISO | ||
| TH1 (cm) | Stand | 0.44 | 0.48 | 0.3 | 0.31 | -0.27 | -0.06 |
| Family | 0.2 | 0.35(.014) | 0.33(.020) | 0.35(.013) | -0.11 | -0.2 | |
| TH2 (cm) | Stand | 0.48 | 0.58 | 0.5 | 0.52 | -0.19 | -0.18 |
| Family | 0.25 | 0.41(.004) | 0.44(.002) | 0.44(.002) | -0.05 | -0.1 | |
| ΔTH (cm) | Stand | 0.02 | 0.14 | 0.38 | 0.41 | 0.21 | -0.27 |
| Family | 0.07 | 0.08 | 0.16 | 0.13 | 0.09 | 0.16 | |
| DB1 (mm) | Stand | 0.26 | 0.29 | 0.3 | 0.14 | -0.02 | 0.58 |
| Family | 0.14 | 0.38(.007) | 0.23 | 0.21 | -0.28 | 0.02 | |
| DB2(mm) | Stand | 0.15 | 0.54 | 0.2 | 0.04 | -0.4 | 0.65 |
| Family | 0.02 | 0.37(.009) | 0.19 | 0.15 | -0.29(.044) | 0.22 | |
| ΔDB (mm) | Stand | -0.12 | 0.37 | -0.12 | -0.13 | -0.52 | 0.15 |
| Family | -0.1 | 0.14 | 0.04 | 0 | -0.12 | 0.28 | |
| DB2/TH2 | Stand | -0.47 | -0.25 | -0.42 | -0.54 | -0.1 | 0.61 |
| Family | -0.30(.038) | -0.24 | -0.41(.004) | -0.44(.002) | -0.14 | 0.29(.042) | |
| NLSprouts | Stand | 0.41 | 0.36 | 0.53 | 0.04 | 0.04 | 0.17 |
| Family | 0.16 | 0.27 | 0.32(.025) | 0.32(.025) | -0.03 | -0.08 | |
| DevVerticality (Degrees) | Stand | 0.21 | 0 | -0.25 | -0.21 | -0.21 | -0.1 |
| Family | 0.14 | 0.16 | 0.17 | 0.15 | -0.06 | 0.11 | |
| NeedLength (mm) | Stand | 0.05 | 0.19 | 0.25 | 0.27 | 0.05 | -0.2 |
| Family | 0.15 | 0.05 | 0.14 | 0.15 | 0.09 | -0.11 | |
| NeedShLength (mm) | Stand | -0.22 | -0.3 | 0.3 | 0.26 | 0.58 | -0.02 |
| Family | 0.04 | -0.01 | 0.23 | 0.21 | 0.24 | 0.06 | |
| NeedShLength/ NeedLength | Stand | -0.22 | -0.38 | 0.25 | 0.18 | 0.61 | 0.14 |
| Family | 0 | 0.01 | 0.18 | 0.15 | 0.17 | 0.16 | |
| NStDS | Stand | 0.01 | 0.28 | 0.04 | 0.11 | -0.27 | -0.1 |
| Family | -0.30(.038) | -0.04 | -0.01 | 0.02 | 0.08 | -0.2 | |
| NStVS | Stand | -0.6 | 0.01 | 0.42 | 0.41 | 0.31 | -0.58 |
| Family | -0.27 | -0.07 | -0.16 | -0.15 | -0.6 | -0.02 | |
| NStM5 | Stand | -0.54 | -0.76(.03) | -0.6 | -0.51 | 0.26 | -0.24 |
| Family | -0.14 | -0.31(.028) | -0.14 | -0.13 | 0.29(.045) | -0.04 | |
NCOT: Number of cotyledons; LHYP: Hypocotyl length; LOCOT: Longest cotyledon; SHOCOT: Shortest cotyledon; COTANISO: Cotyledon anisotropy (LOCOT/SHOCOT). Statistically significant (p<0.05) coefficients are in bold type along with their probabilities in parentheses.
Table 5: Linear Correlation coefficients between Plantlet and Seedling Traits at the Stand (N=8) and the Family-within-Stand levels (N=49) levels.
Correlation among seedling traits and geographic parameters
The coefficients of correlation between seedling traits and longitude, altitude are depicted in Table 6. At the stand level, Slenderness of the seedlings at age 2 (DB2/TH2) and the Number of Stomata on the dorsal side of the needle (NStDS) were the only traits which display significant coefficients of correlation with the Kilometric longitude (r = –0.88, p = 0.04 and r = 0.80, p = 0.02, respectively). There were no traits, at the stand level, correlated to Altitude.
| Trait | Stand level | Family-within-Stand level | ||
|---|---|---|---|---|
| Longitude | Altitude | Longitude | Altitude | |
| TH1 (cm) | 0.33 | 0.04 | 0.36(0.01) | -0.06 |
| TH2 (cm) | 0.67(0.07) | 0.18 | 0.43(0.002) | 0.03 |
| ΔTH (cm) | 0.66(0.07) | 0.34 | 0.1 | 0.14 |
| DB1 (mm) | -0.57 | -0.24 | -0.05 | -0.04 |
| DB2(mm) | -0.35 | 0 | 0.1 | 0.07 |
| ΔDB (mm) | 0.16 | 0.27 | 0.18 | 0.14 |
| DB2/TH2 | -0.88(0.04) | -0.1 | -0.45(0.001) | 0.05 |
| NLSprouts | 0.07 | -0.22 | 0.09 | -0.06 |
| DevVerticality(Degrees) | 0.33 | 0.05 | 0.13 | 0.06 |
| NeedLength (mm) | -0.08 | -0.18 | -0.01 | -0.14 |
| NeedShLength (mm) | -0.42 | -0.35 | -0.22 | -0.12 |
| NeedShLength/NeedLength | -0.43 | -0.38 | -0.19 | -0.03 |
| NStDS | 0.80(0.02) | 0.66 | 0.28 | 0.27 |
| NStVS | 0.21 | 0.47 | 0.08 | 0.22 |
| NStM5 | -0.04 | 0.21 | -0.27 | 0.03 |
Statistically significant coefficients are in bold type followed in brackets by the probability of error.
Table 6: Coefficients of Correlation between seedling Traits at ages 1 and 2 and geographic variables (Kilometric longitude, altitude) at the stand (N=8) and the family-within-stand levels (N=49).
At the family-within-stand level, a significant negative coefficient of correlation is confirmed for Slenderness (r = –0.45, p = 0.001) along with significant positive coefficients for height at ages 1 and 2 (r=0.36, p = 0.01 and r=0.43, p = 0.002, respectively), meaning that the eastern families tend to be more vigorous than the western ones.
The calculated coefficient of conformation between the frequencies of the pre-supposed stand-membership of the families and the frequencies of wrongly-classed families was not statistically significant (C = 0.35) Chi2 = 9.35; df = 7; p = 0.23, see annex). This indicates a lack of structure differentiation between the Aleppo pine stands in this region for the studied traits at one- and two-years old seedlings stages (Figure 3).
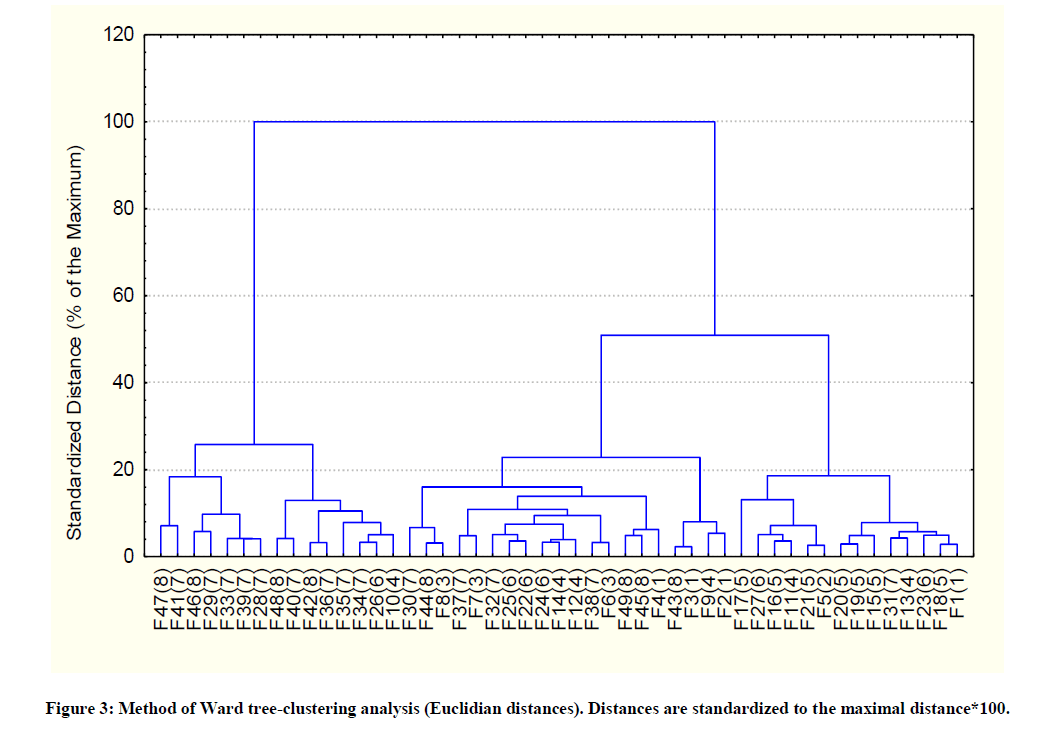
Figure 3: Method of Ward tree-clustering analysis (Euclidian distances). Distances are standardized to the maximal distance*100.
One can observe the large scattering of the families of a same stand denoting a lack of structure differentiation of the Aleppo pine stands for juvenile traits in the studied region. Legend: F47(8) is the 47th family (from the 8th stand).
Under climate changes, Genetics of morphological and growth traits is worthy to know owing to the importance of these characters with regard to adaptation and fitness of individuals, populations and species. Also, seedling characters, since measured in a common environment, allowed us to acquire population structure information and to estimate genetic parameters, and could be highly interesting for early selection purposes. Due to harsh environmental conditions, regenerating forest areas is proving very difficult and often disappointing; so, early selection of seedlings and families for growth is to enhance the capability of seedlings to withstand competition in the field following their transplantation.
Except a slight (p<0.05) significant variation for Total Height and Basal Diameter of the seedlings, there was no large differences among stands at one- and two-years old seedling stages. Throughout the studied region, Analyses of Variance, Coefficients of geographic differentiation and, above all, Hierarchical Classification, showed no substantial structure differentiation between stands for one- or two-years old seedling characters. Within such a small area, gene flow is likely to erase any sort of structure differentiation among stands [10,11]. Indeed, exchange of genetic material by pollen and/or seed between populations is yet largely possible little by little due to the existence of isolated go-between trees. Consequently, scattered stands in this zone are likely not so completely isolated by distance and could share similar pool of genotypes, resulting in a very small differentiation among them. Furthermore, these stands are probably the remnants of a more widespread ancestor population which was formerly split by agriculture practice and, nowadays, relegated to marginal sites for environmental purposes.
On the other hand, family-within-stand was a relatively important source of variation especially for characters linked to growth; although some were statistically significant, morphological and stomata traits were less variable than growth characters. A strong within-stand variation may occur when high level of genetic variance exists in populations or when collected seeds are inbred due to selfing and/or mating between relatives within neighbourhoods or small groups of trees in a population [13-15]. In addition, at such early stages, mother effects due to the nutritive seed tissue, which is a sort of environmental source of variation, could play a role in (non-genetically) increasing resemblance among siblings, especially for growth traits. All of these effects should work together for resulting in overestimating intra-class correlations and heritabilities [14,16]. However, parameters as size and location of stands may play a role in shaping genetic variation within stands. This variation was generally of lesser importance in small isolated and/or coastal stands. Stands of small size may, eventually, differentiate by random drift [17-20]; coastal stands, owing to their more or less linear configuration and the presence of the sea on one side, are also less exposed to receive exogenous genes by migration [21].
Since growth traits are closely related to fitness, high values of narrow-sense heritability are an asset for the practice of early family or individual selection and a guarantee for future adaptation, mostly under global climate change [13,22]. Growing fast in their first and second years of life is a decisive selective advantage for seedlings to withstand severe competition conditions.
Genotypic age-age correlations between traits at the stand level as well as at the family level were relatively weak for most characters but growth (Total height and Basal diameter). Correlations between the same traits at different stages might be used as criteria for juvenile selection at more or less advanced stages.
The correlation between seedling traits and longitude (Kilometric) at the stand level was statistically significant for Slenderness (ratio Basal Diameter/Total Height) and the density of Stomata on the dorsal side of the needle; positive and relatively high, but significant only at 7%, coefficients were observed for Total Height. The significant (negative) coefficient of correlation was confirmed at the family-within-stand level for Slenderness but not for the density of stomata, along with significant positive coefficients for Total Height at ages 1 and 2, meaning that the eastern families tend to be more vigorous than the western ones. The studied zone spans about 15 km west-east and 3 km north-south from the Mediterranean shore; it was not expected to find clines within so a small area. The clines were west-east and parallel with an eastward increase in annual rainfall records (628mm/year vs 717 mm/year, respectively); a positive relationship seems to exist between vigor and precipitation quantities; such a clinal variation in forests stands is frequently recorded for other forest species but other parameters other than rainfall can be involved such as cold, altitude or latitude by their effects on temperatures and precipitations as well as on the growing season duration [18]. In our case, nevertheless, inbreeding effects or drift also cannot be disregarded because of the greater occurrence of small stands in the western part of the zone hence increasing the probability of homogamy or random losses [17,18,21].
Few works are available on phenotypic variation at the within-region-of-provenance scale in Aleppo pine, especially at juvenile stages; extant works focusing mostly on the regional and/or the global area of the species, provenances or regions of provenances.
In Algeria, and elsewhere in North Africa, some studies found a relatively significant phenotypic variation among provenances for morphological and adaptive traits [9,23,24] but a relatively low variation in isozymes [5,11]. The frequency of allele Adh21 in Aleppo pine is low and reaches a maximum (referring to the works made until now on the topic) of 0.11 in the population of Wadi Latrum in the region of Syrte, Libya [12] and a study on 14 moroccan populations [5] showed low allelic richness and heterozygosity using five enzyme systems. Results obtained from these studies reinforce the knowledge that Aleppo pine in North Africa lacks genetic polymorphism for isozyme markers. The low genetic differentiation for such markers as isozymes, less sensitive to natural selection/environmental variations, let us invoke the possible existence of a foundation permanent bottleneck due the recurrence of fires at short intervals of time. In North Africa, fires are an important factor of evolution for forest species, and these species developed various strategies for adapting themselves; Aleppo pine, presents, for example, serotinous fruits and the capacity of producing fertile seeds from an age as young as 7 or even earlier.
Elsewhere, studies of local provenances or from the whole range of the species revealed a significant variation among populations using various genetic markers and traits whose some were very heritable [2][25-31]. In South-Eastern-Australian experiments grouping populations from the whole range of the species, it was observed a significant geographic variability in seedling characters [25]. Generally, wide tests of Aleppo pine provenances highlighted a large geographic variability for morphological and adaptive traits; inversely, it seems that such a high variability do not exist among stands of the same region of provenance as it was recorded in the present work, where seeds were collected from stands separated by small distances up to circa 15 km, However, in other pine species such as jack pine (Pinus banksiana), it was observed a significant differentiation for some seedling traits among stands separated by relatively short distances up to 25 km [32,33-35]. So, such findings could not be generalized for all traits and/or all pine species.
As a direct implication of these findings, it can be sufficient, when prospecting at a local scale, to focus efforts on families and/or individuals, rather than on stands, for selection purposes as long as early selection, for initial growth for example, is aimed. Initial growth during the nursery is worthwhile for the survival of seedlings after transplantation while subjected to competition. On the other hand, the family within-stand variability should be exploited to face climate changes.